Lupine Publishers | LOJ Pharmacology & Clinical Research

Introduction
Since the beginning of civilization, medicinal plants have been an intrinsic component of human life [1]. The conservation of ethnobotanical knowledge as part of living culture and practice between communities and the environment is essential for biodiversity conservation. The information about medicinal plants gains from various medicinal systems such as Unani, Siddha, and Ayurveda [2]. The traditional system of medicine belongs to the traditions of each country and has been passed over from generation to generation. Understanding the dynamics of traditional local knowledge of medicinal plants is important for their medicinal properties is now being developed as a source of scientific research to prove the effect of plants and generate new therapeutic resources. Medicinal plants are considered as a backbone of traditional medicine (WHO) as well as most modern medicine is also derived from medicinal plants i.e. aspirin. The medicinal plant having a rich source of components that can be used to develop and synthesize drugs. About 3.3 billion people in developing countries depend on medicinal plants on a regular basis, WHO estimated that about 80% world population rely on the medicinal plant for their primary health care. Further more, worldwide 42% of 25 top-selling drugs marketed are either directly obtained from natural sources or entities derived from plant products [3]. The quality of traditional medicine is determining its active substances produced by the plant. Andrographis paniculata is one of the important medicinal plants that is utilized throughout the world [4]. A.paniculata is an herbaceous plant of the Acanthaceae family. It is widely distributed in Southeast Asia, India, and tropical as well as in subtropical Asia. A.paniculata is also known as the “King of Bitters” since it has a highly bitter taste in all parts of the plant body [5]. Furthermore, A. paniculata is known as “Kalmegh” in India, “Chuan-Xin-Lian” in China, “Fah Tha Lai” in Thailand, “Hempedu Bumi” in Malaysia, “Senshinren” in Japan, and “green chiretta” in Scandinavian nations [6]. A.paniculata is one of the most widely used plants in Ayurvedic and Unani medicine [4]. Traditionally, A.paniculata was used in the treatment for snakebite, fever, bug bite, diabetes, malaria, and dysentery [7]. Moreover, A.paniculata is also used in the combination with other herbs and health care treatment. It is found that A.paniculata is used in more than half of the herbal formulations commercialized in India for he patic diseases [8]. Many scientific studies also have been reported regarding the medicinal properties possessed by the A.paniculata, most of which are based on traditional knowledge (Table 1). Phytochemical investigations have revealed that A. paniculata contains a wide range of chemicals. In addition, experimental evidence also reported that A.paniculata has a broad spectrum of pharmacological activity including anti-bacterial, antidiarrheal, anti-inflammatory, antiviral, antimalarial, anticancer, antimalarial, hepatoprotective, etc. In this review, we briefly discuss ethnobotanical uses, phytochemistry, and recent scientific finding pharmacological activity of the A.paniculata [6].
Table 1: Taxonomical classification of Androgrphis paniculata.
Botanical Description and Habitat
A.paniculata is native species of India, China, and Taiwan. But it is also found in Southeast Asia, tropical and subtropical Asia as well as few other nations such as Malaysia, Indonesia, Vietnam, Sri Lanka, Laos, Cambodia, Pakistan, Myanmar, and the Caribbean islands [9]. Especially, in India A.paniculata are found in Karnataka, Andhra Pradesh, Tamil nadu, Uttar Pradesh, and Madhya Pradesh. Also cultivated in Assam and West Bengal to some extent. In addition, A.paniculata are found in different habitats including forests, farms, plains, hill slopes, dry and wetlands, and wastelands [10]. A.paniculata is bitter in test, an annual herb that is abundantly branched which grows up to a height of 3.-110 cm in a humid, shady area. It has glabrous leaves that are 8.0 cm long and 2.6 cm wide, little white flowers that are rose-purple or light pink, spots on the petals, and corolla with hairs. The stem was found to be dark green in color, 0.4-1.0 m tall, 2-6 mm in diameter, quadrangular with longitudinal furrows and wings on the angles of the younger part [11] as shown in Figure 1.
Figure 1: Andrographis paniculata morphology.

Traditional Uses of Andrographis Paniculata
A.paniculata play vital importance in the Ayurvedic, Siddha, and traditional medicine systems in India [12]. For centuries, the leaves and roots of A.paniculata have been used to treat a wide range of health problems in Asia and Europe. However, the entire plant is also utilized for specific uses [13]. The plant known as “Kalmegh” in Ayurvedic literature is an essential element in the majority of Ayurvedic remedies and is officially recognized by Ayurvedic pharmacopeia. Moreover, it is used as an aperient, emollient, astringent, anti-inflammatory, diuretic, anthelmintic, carminative, and antipyretic in the Unani system of medicine [14].In India, tribal groups used this herb to cure a number of diseases such as antidote against snake bites, Banded Krait and Russell’s viper, etc. [14]. The tribal of Kheria, Khatra, Moora, and the Santal region of Bankura district, West Bengal, India utilizes an infusion of the entire plant to treat fever [15]. The extracted juice from A.paniculata leaves, alone or combined with cloves, cinnamon, and cardamom is used as a cure for flatulence, loss of appetite, griping, diarrhea in children, and irregular stool. In India during the influenza epidemic in 1919, A.paniculata was shown to be highly effective in reducing the disease progression [16]. It was also utilized by ancient Chine’s physicians to treat inflammatory diseases, colds, laryngitis, and fever, hepatitis, pneumonia, respiratory infections, tonsillitis, sores, pelvic snake bites, herpes zoster and it has been characterized as a cold property herb [13] to remove toxins and body heat. The decoction of fresh leaves of A.paniculata is used as an antihypertensive and antidiabetic in Malaysian folk medicine. Furthermore, it is advised to use it in cases of leprosy, scabies, gonorrhea, boils, chronic and seasonal fevers, and skin eruptions, due to its “blood purifying” purifying properties [4].
Phytochemistry
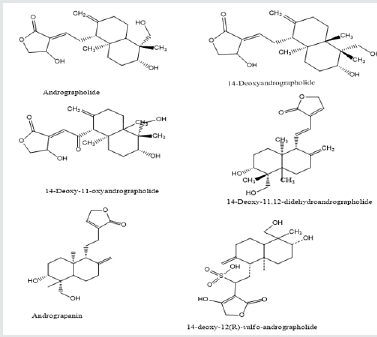
The aerial part (leaves and stems) of A. paniculata contains major active phytochemicals [17]. According to the survey of the literature, andrographolide is the major bioactive compound found in the A.paniculata which is a diterpene lactone that is crystalline, colorless, and has a bitter taste [9]. The leaves have the highest concentration of andrographolide about 2.39% whereas the seed has the lowest concentration about 0.58%. The quantity of the phytochemicals varies widely depending on the portion used, locality, time of harvesting, and season (Figure 2). Andrographolides are highest found immediately before the flowering season, then decline progressively [14]. Other lactones compound observed in A.paniculata is 14-deoxy-11-andrographolide, 14-deoxy-11, 12 didehydroandrographolide, andragraphan, andrographon, 14-deoxyandrographolide, neoandrographolide, deoxyandrographiside, andrographosterol, andrographiside etc. A.paniculata also contains Xanthones and quinic acid derivatives in minor concentrations. Moreover, Reddy et a. [18] reported that A.paniculata contains flavone such as 5-hydroxy-7’2’6’-trimethoxyflavone and 23-C terpenoid 14-deoxy-15-isopropylidene-11, 12- didehydroandrograholide and other flavonoid Skullcapflavone I 2’-O-glucoside, 7-Omethylwogonin, 7-Omethyldihydrowogonin and 7-O-methylwogonin 5-O-glucoside as well as diterpenoids such as isoandrographolide 14-deoxy-11, 12 didehydroandrographolide. Rao et al. [19] identified and isolated 5, 7, 20, 30-tetramethoxyflavanone and 5-hydroxy-7, 20, 30-trimethoxy flavone from the A.paniculata. A new labdane type diterpenoid which is andropanolide along with seven known diterpenoids isolated from the methanolic leaves extract of A.paniculata [20].
Figure 2: Chemical structure of major component found in A.paniculata.
Pharmacological Activity of A.Paniculata
Hepatoprotective activity
A.paniculata is widely used as a hepatoprotective and hepatostimulative agent in the Indian traditional medicine system. Traditionally the leaves aqueous extract of A.paniculata is used in the treatment of jaundice and different liver damage. Andrographolide found in the A.paniculata was protective against liver damage in rats and mice induced by carbon tetrachloride. Moreover, Andrographolide also observed significant hepatoprotective against various types of liver damage, induced by galactosamine or paracetamol [21]. The free radical scavenging activity of andrographolide has a significant hepatoprotective effect by lowering lipid peroxidation malondialdehyde product as well as by maintaining glutamic pyruvate transaminase, alkaline phosphatase, and glutathione levels in mice treated with carbon tetrachloride [22]. A.paniculata has been shown antihepatotoxic activity against plasmodium berghei K173-induced hepatic damage in mastomys natalensis [23].
Antibacterial activity
A.paniculata has been shown the antibacterial activity against a wide range of bacterial species. In vitro study found that the aqueous extract of A.paniculata shown antibacterial activity even at the low concentration (25 mg/ml) against E.coli, Shigella, Streptococci, Staphylococcus aureus, and Salmonella [24]. Another similar study leaves aqueous extract of A.paniculata reported against the methicillin- resistant S.aureus and Pseudomonas aeruginosa [25]. Furthermore, A.paniculata is also effective against HAS 1 (herpes simplex virus 1) without any cytotoxicity [26].
Antidiarrheal activity
In developing countries, Diarrhea is one of the most common diseases and it leads to the top ten causes of death among children worldwide [5]. Some drugs such as kaolin-pectin, selenium, loperamide, and bismuth have been used to treat the symptoms. However, it also causes some unfavorable side effects [5]. The study has been found that A.paniculata has significant antidiarrheal properties [27]. According to the study, an ethanolic extract of A.paniculata treated 88.3 % of acute bacillary dysentery cases and 91.3% of acute gastroenteritis cases. Furthermore, andrographolide was found to treat 91% of acute bacillary dysentery cases. The same cure rate of about 91.1% was obtained by providing a compound tablet comprising andrographolide and neoandrographolide in a 7:3 ratio. This was claimed to be more than the cure rate observed with chloramphenicol and furazolidone [28]. A.paniculata was found to be effective in curing patients with acute diarrhea and bacillary dysentery in double-blind investigation [14].
Antimalarial activity
In many tropic and subtopic countries, malaria is still a prevalent disease [14]. A.paniculata was shown to significantly suppress the growth of the Plasmodium berghei [11]. In vitro study of 50% ethanolic extract of the aerial parts (100 mg/g) shown antimalarial activity against plasmodium berghei and in vivo study in rats observed antimalarial activity after intragastric application (1g/kg body weight) [26]. It is suggested that the antimalarial effect of A. paniculata is due to the reactivation of the enzyme superoxide dismutase [5]. Another study has been reported that the crude extract of A.paniculata shown antimalarial activity against the resistant strain of Plasmodium falciparum having an IC50 value of 6mg/ml [29]. In addition, a xanthous compound isolated from the A. paniculata has been shown in vivo antimalarial activity in plasmodium infected Swiss albino mice. The results found that a significant reduction in parasitemia after treatment with a 30 mg/kg dosage [26].
Anticancer activity
Cancer is a set of disorders characterized by abnormal cell proliferation and the ability to penetrate or be spared to other regions of the body. Despite the fact that many diseases have a worse prognosis than most cancers [17]. The extract of A.paniculata having diterpenoid is significantly able to restrict cell proliferation, arrest the cell cycle and induce cell apoptosis of different cancer cells [30- 33]. Treatment of the MDA-MB-231 breast cancer cells with andrographolide extracted from A. Paniculata causes apoptosis of cancer cells and arrests the cell cycle without interfering with the normal growth of cells [34]. The study has been reported that A.paniculata exhibits potent cytotoxic activity against human epidermoid carcinoma of the skin lining of the lymphocytic leukemia cells and nasopharynx [12]. A.paniculata also shown cytotoxic effects against colon cancer cells by suppressing AKT and mTOR phosphorylation levels, resulting in ER stress-induced death [35]. Furthermore, apoptosis in colon cancer cells is induced by the andrographolide via controlling the signaling of pro-apoptotic GRP-78/IRE1/XBP-1/ CHOP [17].
Antidiabetic activity
Diabetes is a metabolic disease characterized by elevated blood sugar levels [36]. According to the WHO reports around 70 million people worldwide suffer from diabetes. Specifically in developing countries, diabetes has become a threat to human health [37]. In vivo study observed that ethanolic extract of A.paniculata exhibit the protective effect in hyperglycemic condition and also protect the tissue damage caused due to oxidative stress in a diabetic rat model produced by streptozotocin [38]. Another study conducted [39] found that oral administration of andrographolide in a dose-dependent manner reduced plasma glucose levels in diabetic rats caused by streptozotocin and wild-type rats.
Conclusion
The entire literature review indicated that Andrographis paniculata exhibits a broad range of phytochemicals and pharmacological activities. The previous study found that A.paniculata contains 50 lactane diterpenoids, 30 flavonoids, and 30 novels phytochemical isolated and identified from A.paniculata. Phytochemical study reveals that Andrographolide is a major compound found in Andrographis paniculata. It has shown a wide spectrum of pharmaceutical activity such as anti-microbial, hepatoprotective activity, anti-inflammatory activity, anti-malarial, anti-diarrheal, anti-diabetic, and cytotoxic activity. The precise information offered as a review here covers the phytochemical and pharmacological information about this plant, providing the muchneeded encouragement to use this plant in creating and sustaining a prospective means of livelihood.
For more LOJ Pharmacology & Clinical Research Journals please click on below link
https://lupinepublishers.com/pharmacology-clinical-research-journal/
